
Annex "Beaches and dunes"
Annex 1. Notes on the vegetation maps and their comparison
The mapping surveys within the trilateral Wadden Sea follow different methods and not all adhere to the method as described in the TMAP handbook. Therefore, some background information is given on the data used in this chapter.
As trilateral harmonisation is in progress and mapping requirements within countries change, mapping methods and typologies within the regions have changed over time. Consequently, trend analysis is not possible except for Hamburg, and no trends have been tried to derive in this QSR. It is expected that this will be possible in the next QSR, as mapping efforts that are underway in Germany allow comparison with the currently reported maps.
Data sources
- The Netherlands: 2005 – 2015. The data are not harmonised between mapping organisations and large dune areas (mostly woodland) are not mapped. For Texel, no new maps have been produced since the QSR 2009. Not all types of the used local typologies can be unequivocally translated into TMAP types without local knowledge. Beaches are not mapped. Harmonisation is underway between some of the authorities.
- Texel and Vlieland: Staatsbosbeheer, Rijkswaterstaat, Ministerie van Defensie
- Terschelling and Ameland: Staatsbosbeheer, Rijkswaterstaat
- Schiermonnikoog: Rijkswaterstaat, Natuurmonumenten
- Richel: IMARES
- Griend, Rottumerplaat, Rottumeroog and Zuiderduin: Rijkswaterstaat
- Lower Saxony: No new maps have been produced since the QSR 2009.
- Nationalparkverwaltung Niedersächsisches Wattenmeer and NLKWN (together): 2004.
- Hamburg: Nationalparkverwaltung Hamburgisches Wattenmeer (Behörde für Umwelt und Energie Hamburg): 2014.
- Schleswig-Holstein: Nationalparkverwaltung Schleswig-Holsteinisches Wattenmeer / Landesbetrieb für Küstenschutz, Nationalpark und Meeresschutz Schleswig-Holstein: 2011-2012.
- Denmark: Miljø- og Fødevareministeriet: 2010 – 2012.
Web portals for viewing vegetation maps are:
- Netherlands Rijkswaterstaat
- Lower Saxony
- Denmark (to view Habitat maps: DEVANO kortlaegning)
Notes to the analysis
- In contrast to the QSR 2004, areas that were not mapped are not included (i.e. they are not reconstructed from other maps).
- In Lower Saxony, X.2 and X.3 are together mapped as one unit, and X.4 and X.5 as another separate unit.
- For the analyses, all types are included, except for X.1 as that is unvegetated beach plain.
- Several types of open dunes and dunes slacks (i.e. H.7, X.9 and X.12) are included, as they are clearly part of the dune landscape.
- X.2 is included for the regions where it is mapped, even if the area may vary considerably from year to year.
- Green beaches are not mapped as an entity anymore, in contrast to the QSR 2004. The reason for this is that they can only be mapped on a landscape level and not on a vegetation type or habitat type level, which limits the reliability of delineating them. Also, in several cases it is hard to distinguish between salt marsh and green beach, for example on small islands such as Nigehörn. Therefore, total surface areas for vegetation types are reported, dune types (X and H series in TMAP) in this chapter and salt-marsh types (S series) in that chapter .
- The dune types of the xeroseries (the X-series; dry dune vegetation) and hygroseries (H-series; wet dune slacks) correspond –with the addition of two types– to the typology presented in the QSR 2004 (Petersen & Lammerts, 2005).
Annex 2. Dune and beach dynamics per island
Annex 3. Dynamics in established dunes: blowouts
One of the features of dynamic dunes consists of blowouts: unvegetated hollows in which sand is transported. They occur in the foredunes as well as in the more stabilised dunes and can locally rejuvenate the vegetation. Here, we present an example of how such small-scale dynamics can be mapped: for the Dutch Wadden Sea, an overview was made of the number of blowouts per island in 2000 and 2014. The blowouts were mapped from aerial photographs, where shifting embryonic and recently developed white dunes were not taken into account.
The number of blowouts has increased on Texel, but decreased on the other islands, making the total number of blowouts decrease between 2000 and 2014 by ten percent. Vlieland showed the largest relative decline in blowouts. The data show that there are changes in small-scale dynamics in the dunes that could be monitored trilaterally and be related to for example management interventions, coastline development and total dune area. The data are derived from aerial photographs, so that also imagery from the past can be analysed to obtain a larger data series.
There are regionally different views towards the desirability of increased dune dynamics. In The Netherlands, fixed dunes are considered a problem and on several islands active restoration of dune dynamics is carried out by removing vegetation and creating small gaps in foredune. In Lower Saxony, locally sod-cutting is carried out. Norderney represents a special case where the large amount of rabbits contributes to creating blowouts. The dune islands of Hamburg are very dynamic by nature. In Schleswig-Holstein, the traditionally mobile dune areas of Sylt show increased stabilisation, which may lead to a loss of dynamics. In Denmark, a possible loss of dynamics is not a focus for management.
 Figure 51. Number of blowouts in established dunes on the Dutch islands as a measure of dune dynamics (data from the project “Herstel grijze duinen door kleinschalige dynamiek”, funded by O+BN).
Figure 51. Number of blowouts in established dunes on the Dutch islands as a measure of dune dynamics (data from the project “Herstel grijze duinen door kleinschalige dynamiek”, funded by O+BN).
Annex 4. Dune vegetation per island
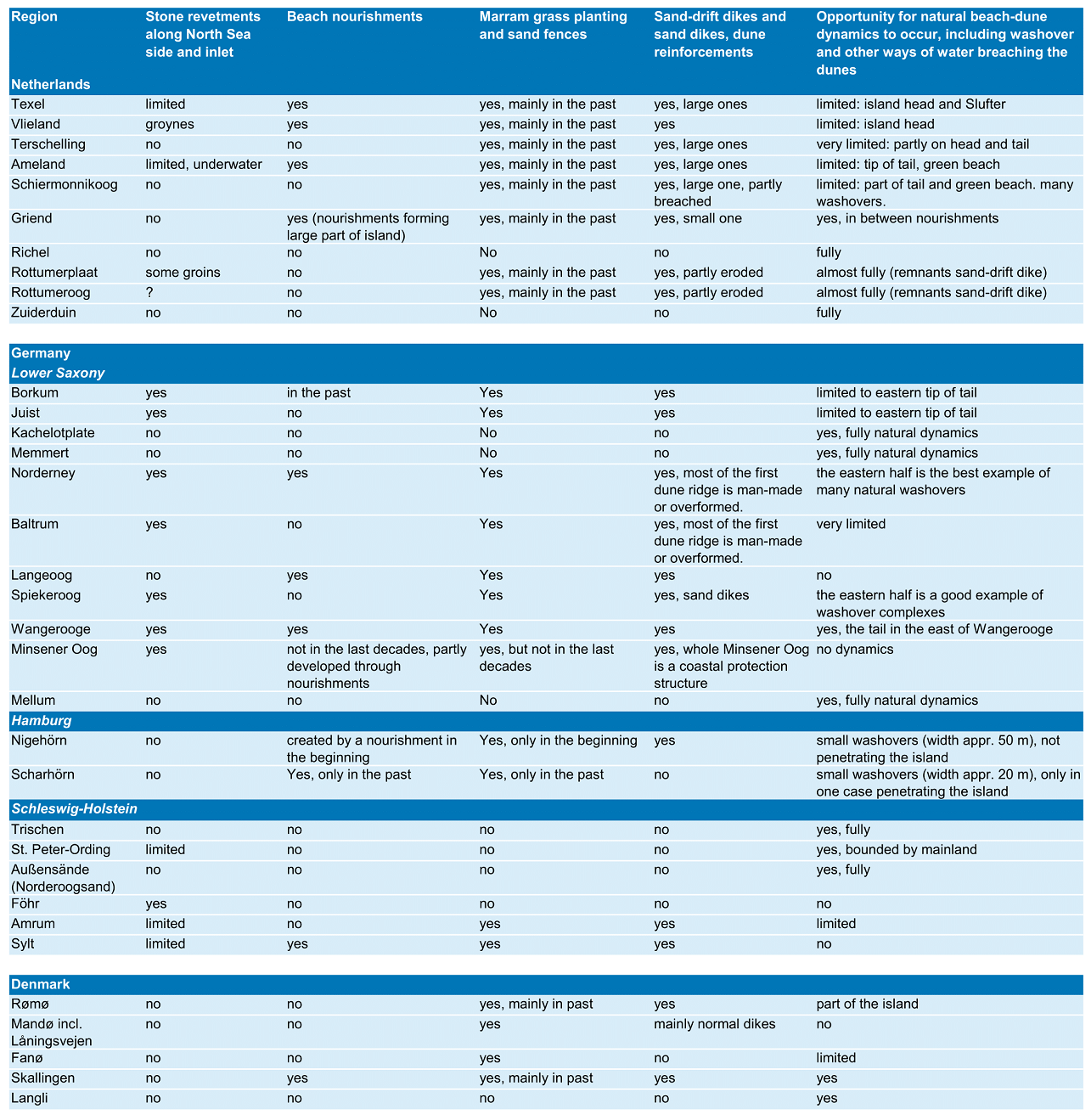
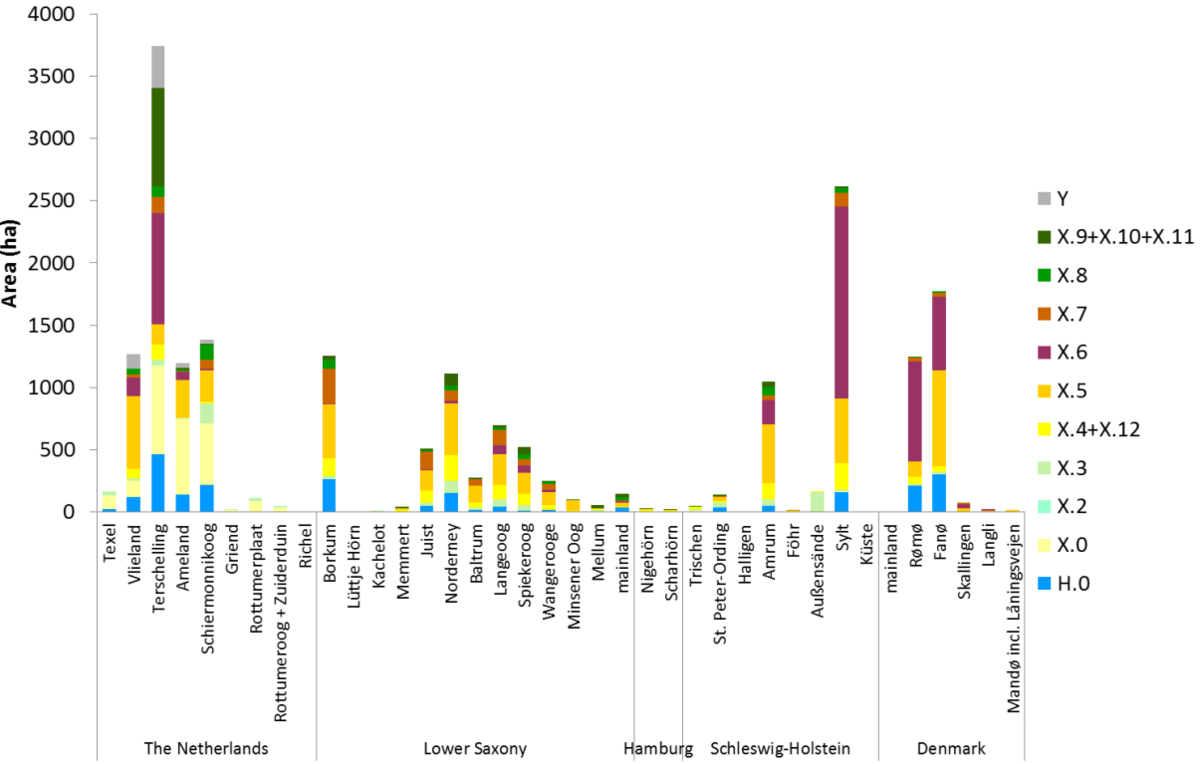 Figure 52. Area of all main dune vegetation types per island. See Petersen et al. (2014) for an explanation of the codes: http://www.waddensea-secretariat.org/management/publications/tmap-typology-of-coastal-vegetation-in-the-wadden-sea-area
Figure 52. Area of all main dune vegetation types per island. See Petersen et al. (2014) for an explanation of the codes: http://www.waddensea-secretariat.org/management/publications/tmap-typology-of-coastal-vegetation-in-the-wadden-sea-area
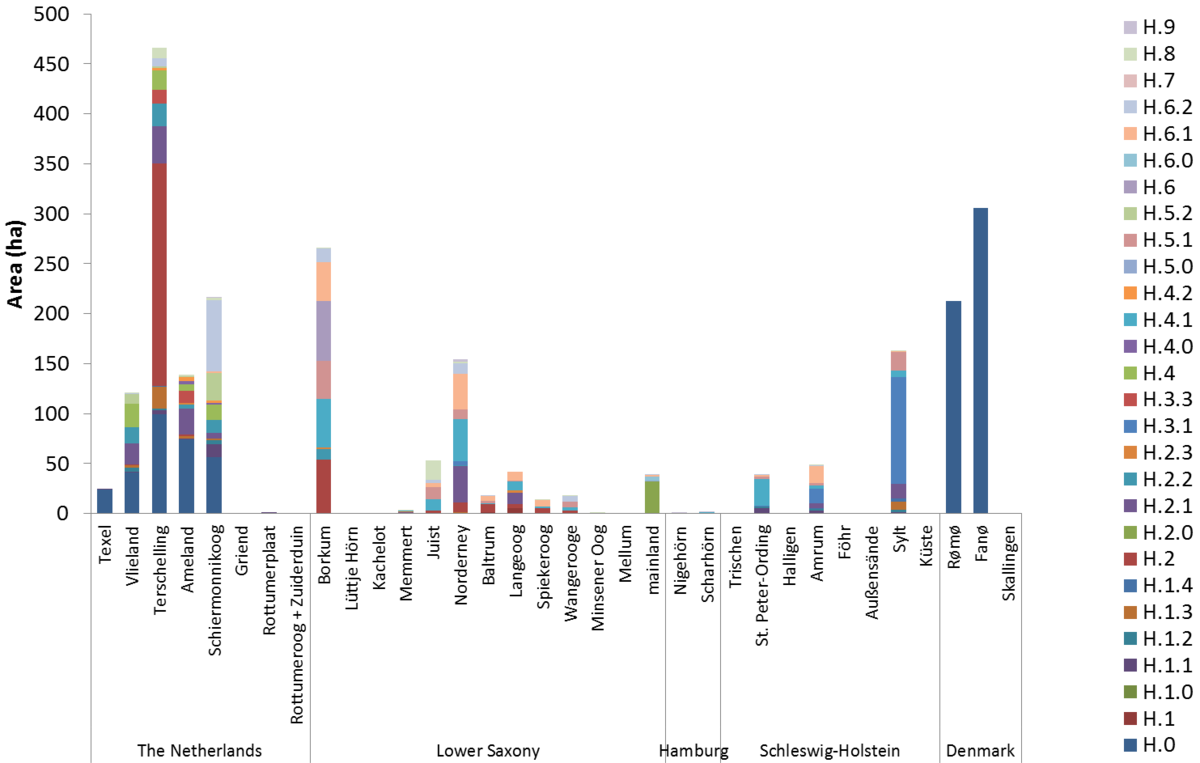 Figure 53. Area of dune slack types per island. See Petersen et al. (2014) for an explanation of the codes: http://www.waddensea-secretariat.org/management/publications/tmap-typology-of-coastal-vegetation-in-the-wadden-sea-area
Figure 53. Area of dune slack types per island. See Petersen et al. (2014) for an explanation of the codes: http://www.waddensea-secretariat.org/management/publications/tmap-typology-of-coastal-vegetation-in-the-wadden-sea-area
Table 4. Area (ha) per island/location.
Will be dowloadable as xls file
Annex 5. Invasive plant species (details)
The moss Campylopus introflexus (Heath star moss) can become dominant very quickly in dry, open short grasslands, such as the Corynephorus canescens (X.5.1) type (Ketner-Oostra & Sýkora, 2004). The dominance of the moss often leads to a decline in bryophyte and lichen richness, but also dune-specific herbs like Jasione montana and Viola canina decline (Van der Meulen et al., 1987; Ketner-Oostra, 1993; Biermann & Daniëls, 1995).
Rosa rugosa (Japanese rose) is particularly invasive in dune grasslands (type X.5), but is able to establish and to dominate in all vegetation types of the xeroseries in dunes (Isermann, 2008). During the die-back of Hippophae rhamnoides bushes (type X.7.1), the release of nitrogen in the soil probably facilitates the establishment and dominance of Rosa rugosa. Normal further vegetation succession is strongly reduced or stopped (Thiele et al., 2010). Rosa rugosa has been planted as an ornamental plant in former times and it often shows increased abundances in close proximity to built-up areas.
Prunus serotina (Rum Cherry) spreads in scattered way in dune grasslands and dune heathlands where organic matter has accumulated. These trees appear to replace indigenous shrubs and trees by shading and halting rejuvenation of native shrub and tree species (Rode et al., 2002; Petersen & Lammerts, 2005; Chabrerie et al., 2010).
Populus alba (White poplar), is considered as alien species in some regions and is invasive in the entire Wadden Sea area. There is only little knowledge about the distribution of other locally invasive Poplar species, like Populus x canadensis.
Senecio inaequidens (Narrow-leaved Ragwort) has been spreading rapidly during the last years (Kuhbier, 1977). It thrives in white dunes (X.4.1 Ammophila arenaria type), dune grasslands (X.5) as well as in Sea Buckthorn shrubland (X.7.1 Hippophae rhamnoides type) and eutrophic dunes (X.10). Abundance in permanent plots is very variable per year, but sometimes the species can become dominant. The species is, as all Senecio species, considered to be toxic.
Pine species like Pinus nigra and Pinus mugo have dispersed considerably along the borders of the original plantations (QSR 2004). The cover degree of the naturalised Pinus species thus decrease with increasing distance to the original plantation (Natter, 2011).
Further invasive alien plant species, mostly in the dunes of the xeroseries, are:
- The garden plants Cerastium tomentosum (Snow-in-summer) and Sedum spurium (Caucasian-stonecrop) are currently naturalised (i.e. established) in dune areas. Fallopia species (F. japonica (Japanese Knotweed), F. sachalinensis (Giant Knotweed), F. x bohemica), Heracleum mantegazzianum (Giant Hogweed), Lobularia maritima (Sweet Alison), and Lupinus polyphyllus (Garden lupin) occur mainly in ruderal dune areas. Whether they have invaded more natural dune sites is yet unknown.
- Claytonia perfoliata (Springbeauty) might become invasive and outcompete small growing scrubland species (Nehring et al., 2013).
- In dune scrubs, Parthenocissus inserta (False Virginia-creeper) is locally invasive.
- Trees and shrubs like Ailanthus altissima (Tree-of-heaven), Amelanchier species (Juneberry), Cotoneaster species, Robinia pseudoacacia (False-acacia), and Spiraea species (Brideworts) are already naturalised in some areas, most of them known to be invasive or potentially invasive (Nehring et al., 2013). In Dutch dunes, Mahonia aquifolium (Oregon-grape), and regional Symphoricarpos albus (Snowberry) are already invasive.
- The liverwort Lophocolea semiteres (Southern crestwort) occurs in dune heathlands, but it is probably often overlooked, so that its distribution is not known well.
In dune slacks, Crassula helmsii (New Zealand Pigmyweed) and in dune ponds Elodea canadensis (Canadian waterweed) might become considerable problems. How invasive Hydrocotyle ranunculoides (Floating pennywort) and Vaccinium atlanticum in dune slacks will be, is not known yet. Vaccinium macrocarpon (Oxycoccus macrocarpos, Cranberry) can already be characterised as invasive species in dune slacks and occurs very frequently on several Dutch islands. A commercial cranberry-culture exists on Terschelling. Locally, some old dune slacks are specifically managed for cranberries by removing succeeding species mechanically and maintaining a high water level by irrigation (Petersen & Lammerts, 2005). The cranberry reaches dominance in older successional stages of dune slacks where once the species-rich Schoenus nigricans type (H.2.2) was present and now has lost its vitality. The cranberry may intrude into the Carex trinervis type (H.2.1) and the Erica tetralix type (H.3.1) and replace native vegetation here (Petersen, 2000).